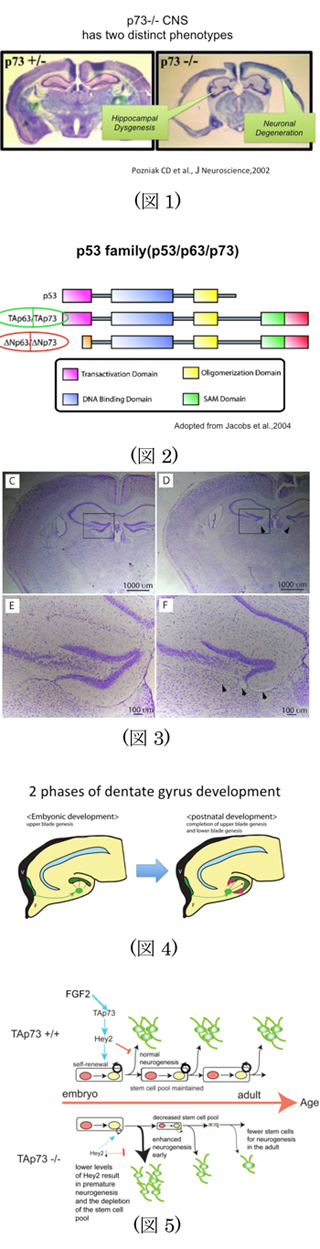
p53ファミリー分子の一つである、p73はそのノックアウトマウスによる解析から、
①皮質の神経変性(細胞死)
②海馬の発生異常
が認められ、脳発生の過程において、また神経細胞の恒常性維持における最重要な分子の一つである。
(Pozniak et al.,science 2000)(図1)
正常発生のみならず、損傷等の病態下におけるその神経保護作用に関して多くの報告がなされてきた。
p73は一つの遺伝子から2つのプロモーターを介して全長型(TAp73)および、アミノ末端欠失型(ΔNp73)二つのアイソフォームが転写され、神経細胞死においては、相互的に拮抗して細胞の生死のバランス制御を行うと考えられてきた。(図2)
しかし、動物個体内でのアイソフォームによる役割分担は、その存在が想定されていたものの、長らく謎であった。
その後、自身が参画したアイソフォーム特異的ノックアウトマウスの解析により、TAp73のノックアウトマウスは成体神経幹細胞の主要な神経新生の場所である海馬の歯状回の異常を来すことを共著者として報告した。
(Tomasini R. Genes and Development 2008
(図3))
冠状断で歯状回を観察すると、歯状回の顆粒細胞層は、神経細胞体を染色する、Nissl染色で示されるとおり、V字型もしくは、U字型をしている。上側をupper blade,下側をlower bladeと便宜上呼称するが、TAp73ノックアウトマウスでは、下側のlower bladeの発生が途中でなくなっていることが分かった。(図3、矢頭)
また、歯状回の顆粒細胞層は、生直後にかけてupper bladeが発生し生後に、lower bladeの発生及び、upper bladeの延長が起こる。(図4)
従って、lower bladeができないということは、出産直後に歯状回に放射線を照射して神経幹細胞を除去した場合見られる表現型と同様であり(Czurko A. et al.,PNAS 1996) TAp73がないことによる神経幹細胞、前駆細胞の枯渇ではないかと考えられた。
そして、TAp73が神経幹細胞の長期的な未分化性の維持を促進することにより、正常な成体神経新生を維持することを証明した。また、TAp73はFGF-2の下流で、bHLH遺伝子であるHey2の転写活性を制御することにより、成体神経幹細胞の早期分化を抑制することで維持する分子メカニズムを証明した。(図5) |